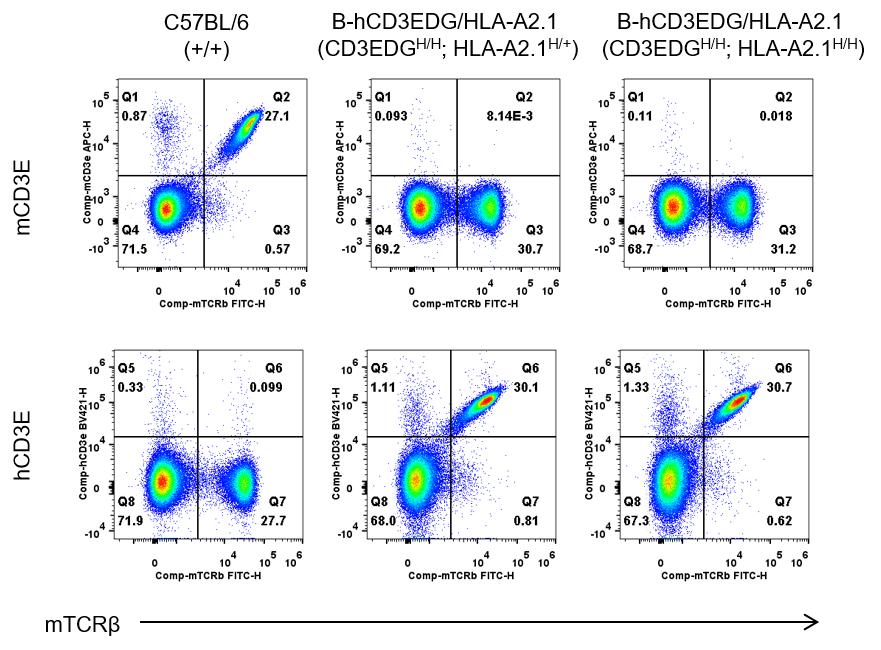
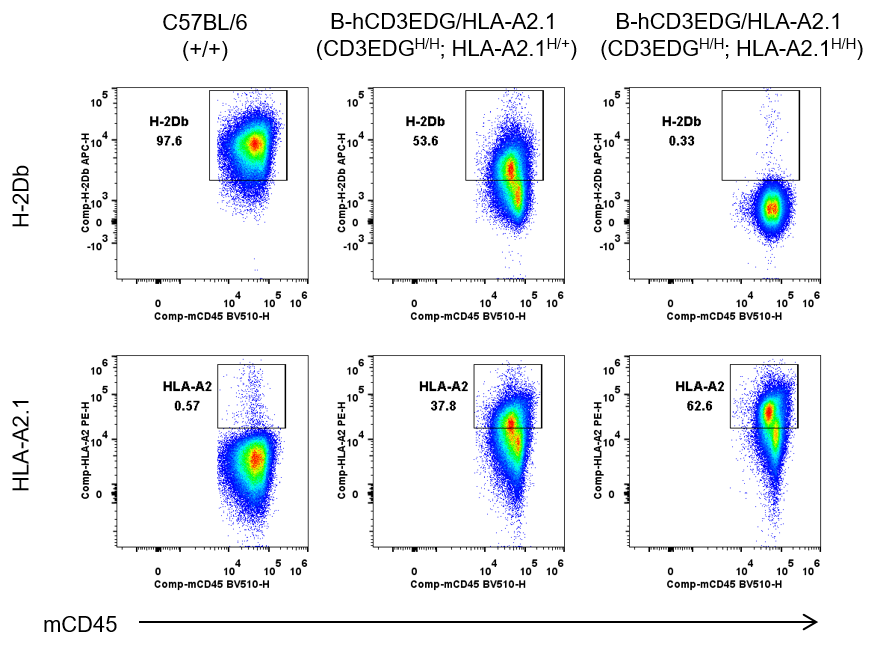
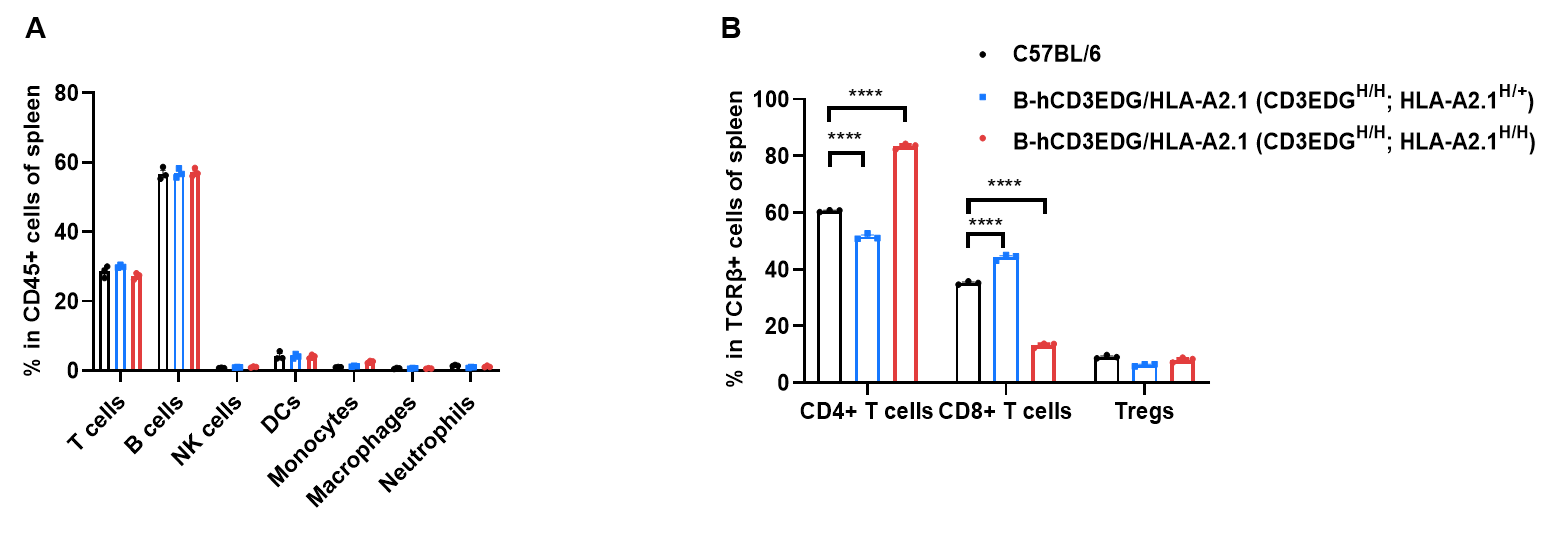
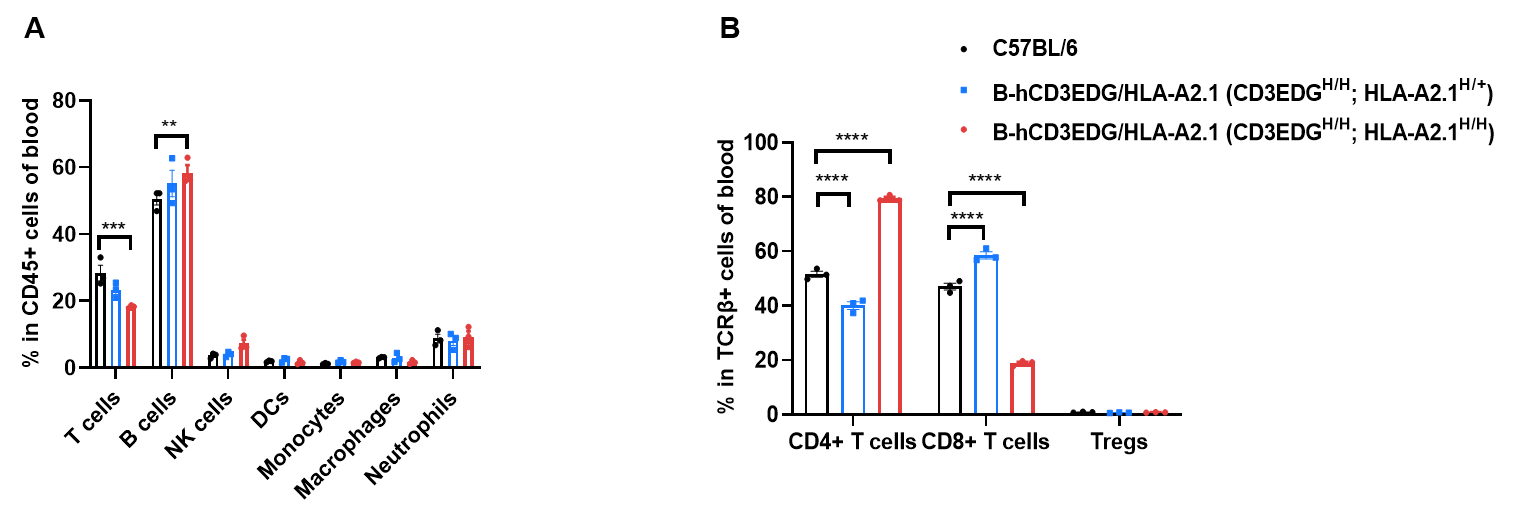
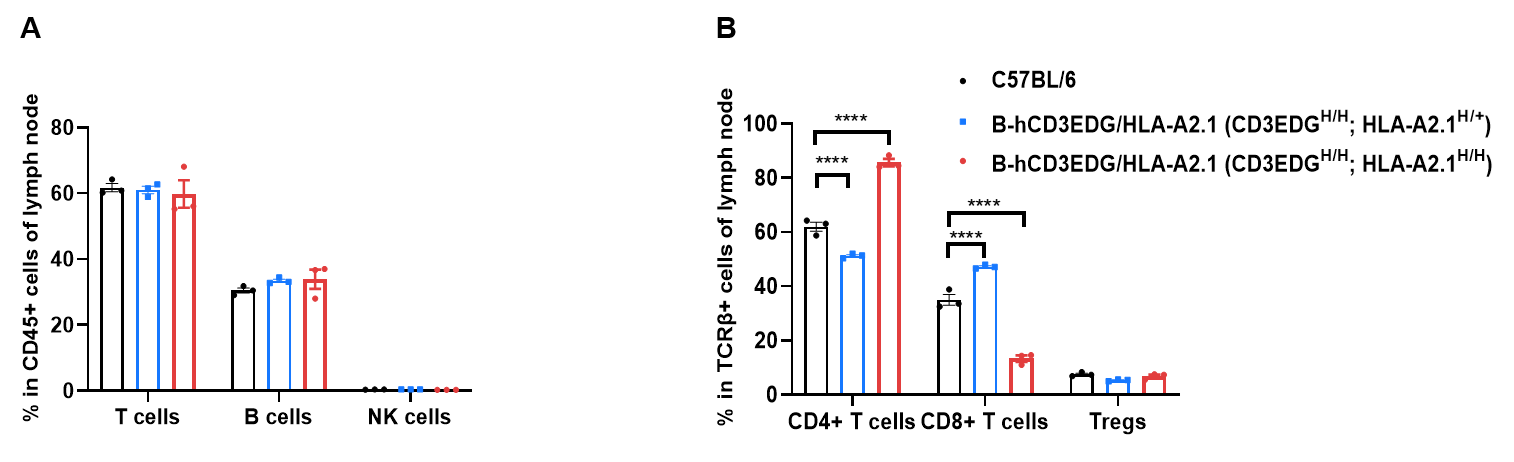
B-hCD3EDG/HLA-A2.1 mice
Catalog Number: 112756
Strain Name: C57BL/6-Cd3etm1(CD3E)Bcgen Cd3dtm1(CD3D)Bcgen Cd3gtm1(CD3G)Bcgen B2mtm2(B2M/HLA-A2.1/H2-D)Bcgen /Bcgen
Strain Background: C57BL/6
NCBI gene ID: 916,915,917,3105 (Human)
Aliases: T3E; TCRE; IMD18; CD3epsilon; T3G; IMD17; CD3GAMMA; CD3-GAMMA; HLAA
---
ライセンスオプション提供可能