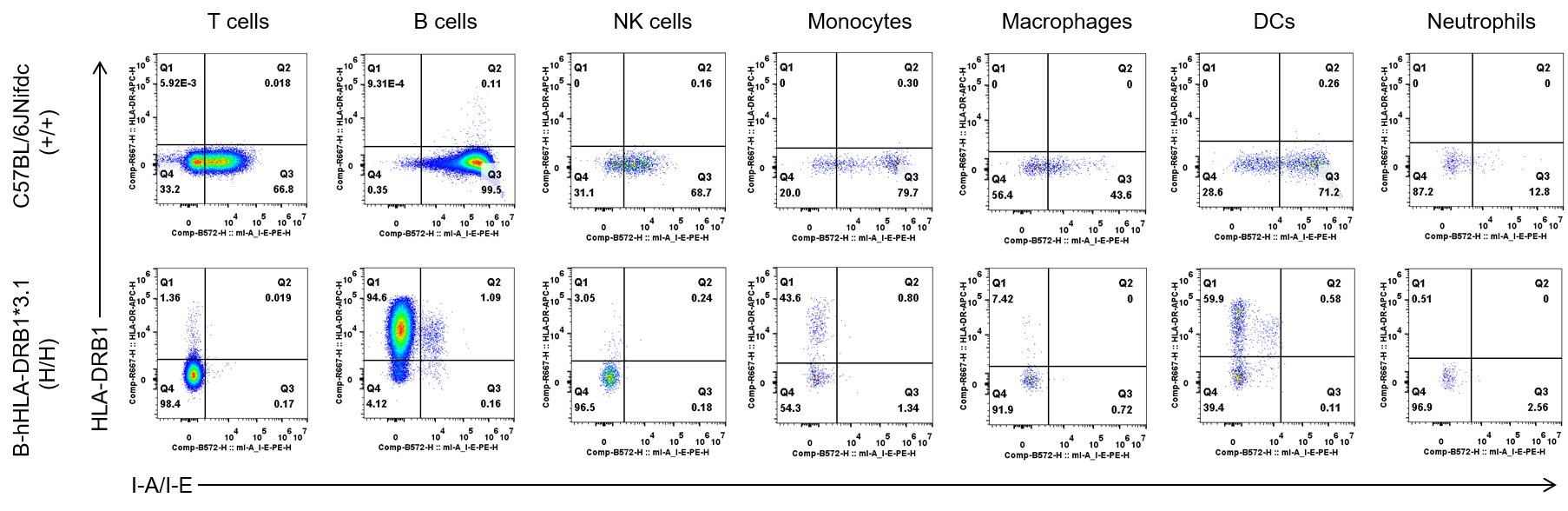
Protein Expression Analysis in Spleen
Strain specific HLA-DRB1 expression analysis in wild-type C57BL/6JNifdc mice and homozygous B-hHLA-DRB1*3.1 mice by flow cytometry. Splenocytes were collected from wild-type C57BL/6JNifdc (+/+) and homozygous humanized B-hHLA-DRB1*3.1 mice (H/H), respectively, and analyzed by flow cytometry with species-specific anti-mouse I-A/I-E antibody (Biolegend, 107607), and species-specific anti-human HLA-DR antibody (Biolegend, 307610). Human HLA-DRB1 was exclusively detectable in homozygous B-hHLA-DRB1*3.1 mice, but not in wild-type C57BL/6JNifdc mice.
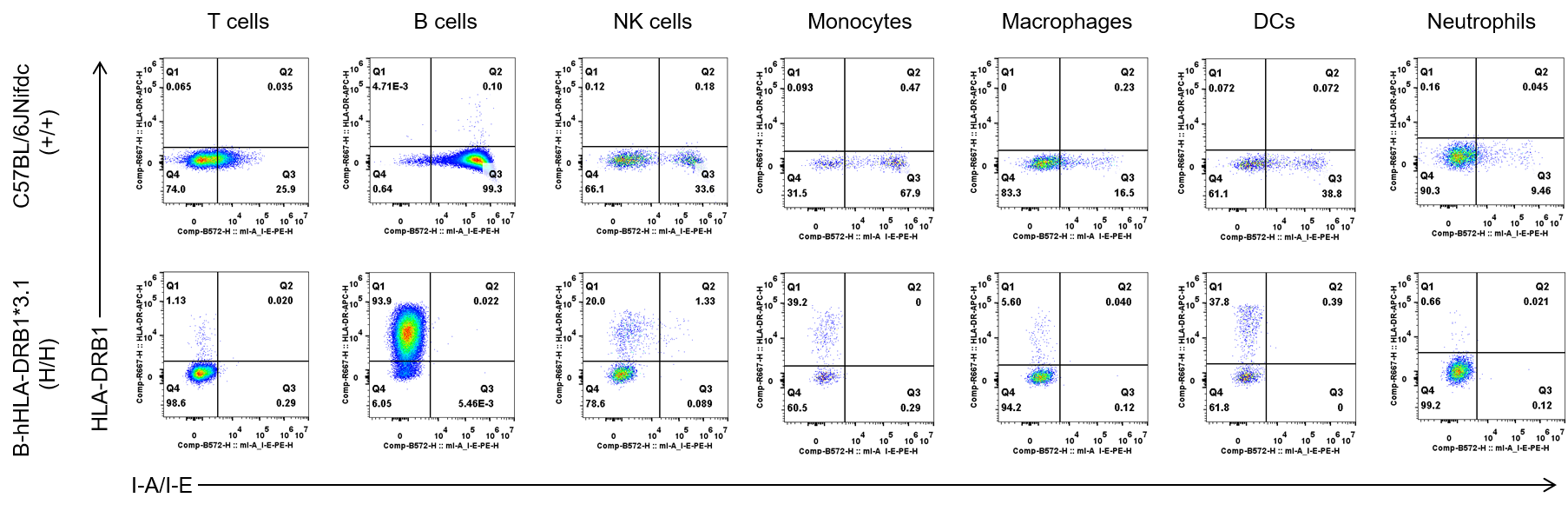
Protein Expression Analysis in Blood
Strain specific HLA-DRB1 expression analysis in wild-type C57BL/6JNifdc mice and homozygous B-hHLA-DRB1*3.1 mice by flow cytometry. Blood cells were collected from wild-type C57BL/6JNifdc mice (+/+) and homozygous humanized B-hHLA-DRB1*3.1 mice (H/H), respectively, and analyzed by flow cytometry with species-specific anti-mouse I-A/I-E antibody (Biolegend, 107607), and species-specific anti-human HLA-DR antibody (Biolegend, 307610). Human HLA-DRB1 was exclusively detectable in homozygous B-hHLA-DRB1*3.1 mice, but not in wild-type C57BL/6JNifdc mice.
Protein Expression Analysis in Bone Marrow
Strain specific HLA-DRB1 expression analysis in wild-type C57BL/6JNifdc mice and homozygous B-hHLA-DRB1*3.1 mice by flow cytometry. Bone marrow cells were collected from wild-type C57BL/6JNifdc mice (+/+) and homozygous humanized B-hHLA-DRB1*3.1 mice (H/H), respectively, and analyzed by flow cytometry with species-specific anti-mouse I-A/I-E antibody (Biolegend, 107607), and species-specific anti-human HLA-DR antibody (Biolegend, 307610). Human HLA-DRB1 was exclusively detectable in homozygous B-hHLA-DRB1*3.1 mice, but not in wild-type C57BL/6JNifdc mice.
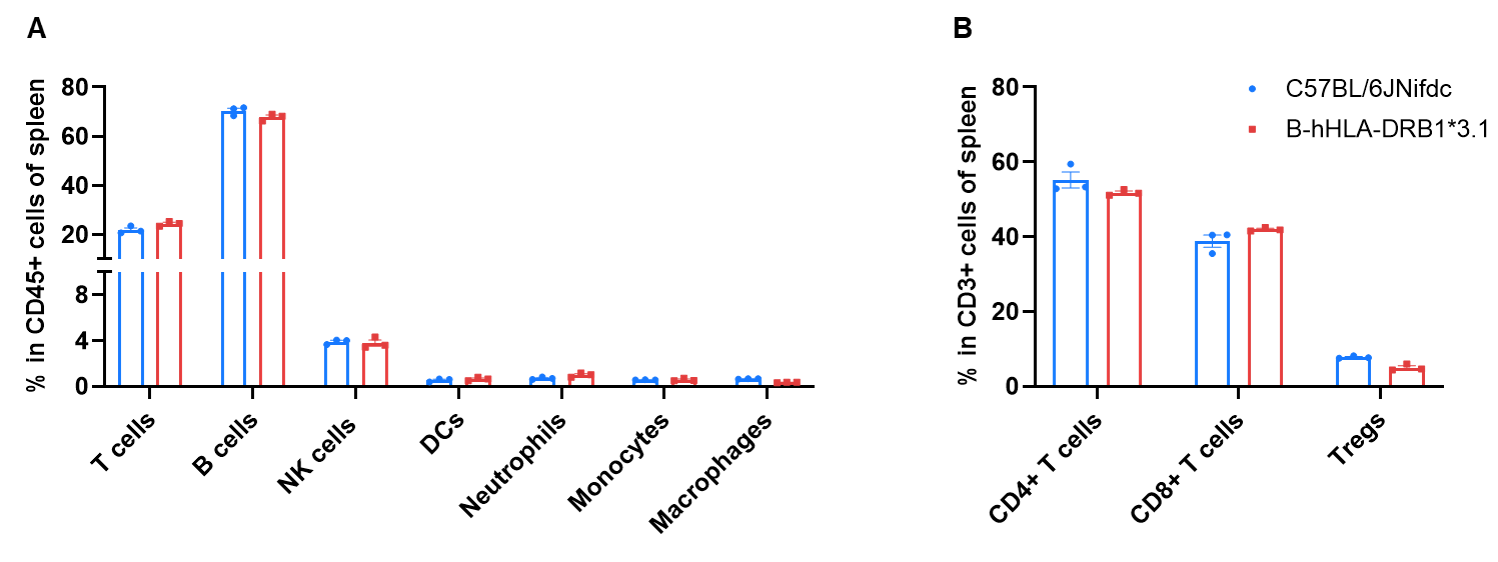
Frequency of Leukocyte Subpopulations in Spleen
Frequency of leukocyte subpopulations in spleen by flow cytometry. Splenocytes were isolated from wild-type C57BL/6JNifdc mice (female, n=3, 8-week-old) and homozygous B-hHLA-DRB1*3.1 mice (female, n=3, 8-week-old). A. Flow cytometry analysis of the splenocytes was performed to assess the frequency of leukocyte subpopulations. B. Frequency of T cell subpopulations. Frequencies of T cells, B cells, NK cells, dendritic cells, neutrophils, monocytes, macrophages, and Tregs in B-hHLA-DRB1*3.1 mice were similar to those in C57BL/6JNifdc mice.
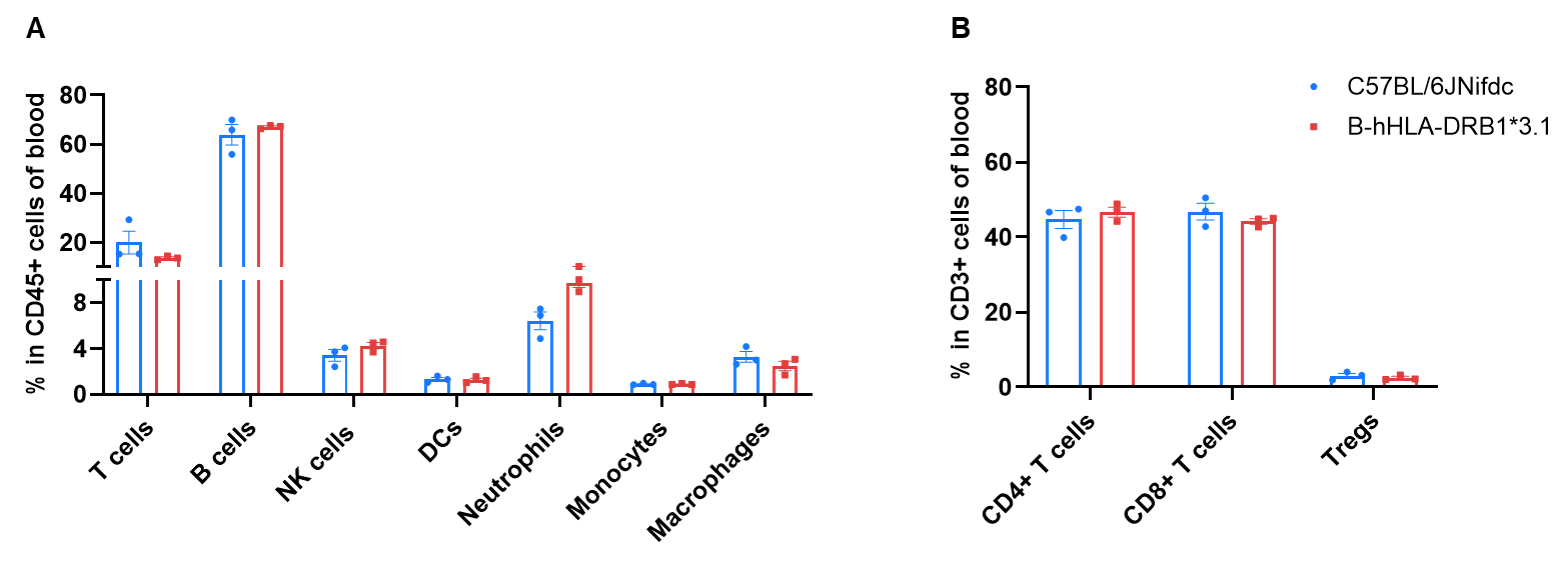
Frequency of Leukocyte Subpopulations in Blood
Frequency of leukocyte subpopulations in blood by flow cytometry. Blood cells were isolated from wild-type C57BL/6JNifdc mice (female, n=3, 8-week-old) and homozygous B-hHLA-DRB1*3.1 mice (female, n=3, 8-week-old). A. Flow cytometry analysis of the blood leukocytes was performed to assess the frequency of leukocyte subpopulations. B. Frequency of T cell subpopulations. Frequencies of T cells, B cells, NK cells, dendritic cells, neutrophils, monocytes, macrophages, and Tregs in B-hHLA-DRB1*3.1 mice were similar to those in C57BL/6JNifdc mice.
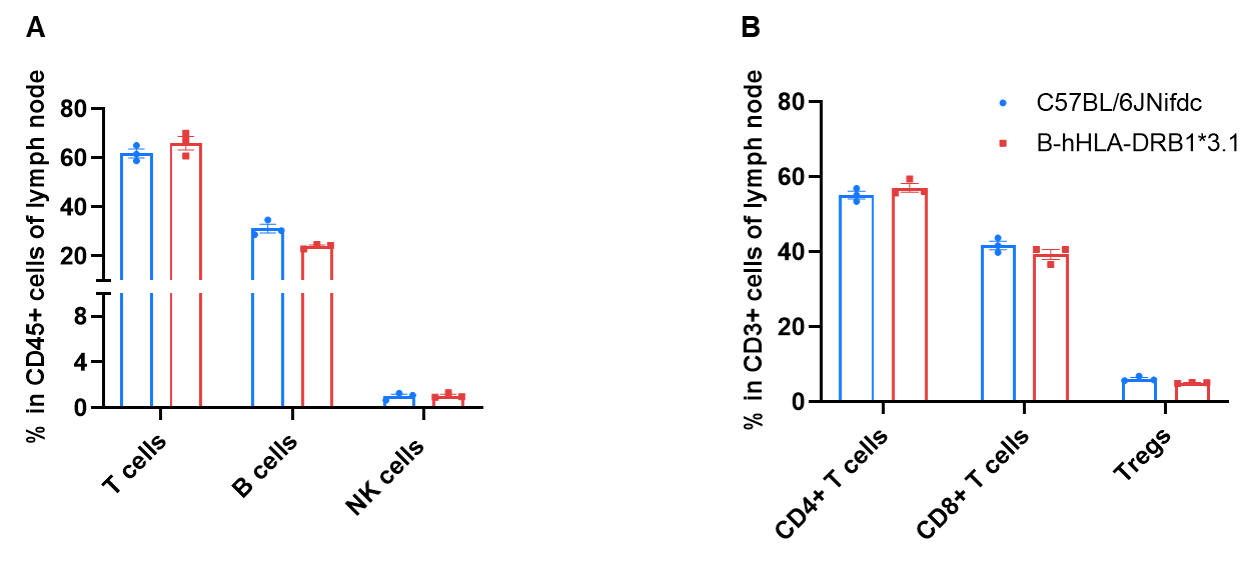
Frequency of Leukocyte Subpopulations in Lymph Node
Frequency of leukocyte subpopulations in lymph node by flow cytometry. Lymph node cells were isolated from wild-type C57BL/6JNifdc mice (female, n=3, 8-week-old) and homozygous B-hHLA-DRB1*3.1 mice (female, n=3, 8-week-old). A. Flow cytometry analysis of the leukocytes was performed to assess the frequency of leukocyte subpopulations. B. Frequency of T cell subpopulations. Frequencies of T cells, B cells, NK cells, dendritic cells, neutrophils, monocytes, macrophages, and Tregs in B-hHLA-DRB1*3.1 mice were similar to those in C57BL/6JNifdc mice.
* When publishing results obtained using this animal model, please acknowledge the source as follows: The animal model [B-HLA-DRB1*3.1 mice] (Cat# 112947) was purchased from Biocytogen.